Nos méthodes
Taxonomie et identification
La taxonomie est une classification du monde vivant basée sur des critères de similitude qui a pour objet de décrire les organismes et de les regrouper. Initialement basée sur la comparaison morphologique, elle s‘appuie désormais sur la biologie moléculaire et tous les outils issus de la génétique.
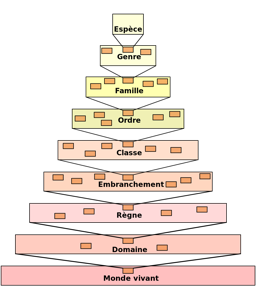
La taxonomie prend une place fondamentale dans nos recherches. La délimitation des espèces, et plus exactement des taxons, se situe à la base de tous les traitements et analyses. La taxonomie évolue perpétuellement, ce qui modifie le regroupement de nos individus. Ainsi un inventaire qui contient 100 espèces à un instant donné sera plus ou moins riche lors de sa relecture en fonction du remaniement taxonomique fait par les taxonomistes. Les espèces, les genres ou encore les familles sont concernés par ces remaniements qui sont publiés dans des revues internationales. Pour assurer la cohérence de nos travaux, nous suivons l’évolution de cette taxonomie au travers du groupe Florical qui publie régulièrement les derniers états de la classification de la flore de Nouvelle-Calédonie en conformité avec la classification mondiale des angiospermes APG IV, proposée par "l’Angiosperm Phylogeny Group".
Identification des plantes
La détermination du nom d’un arbre est une expertise qui repose sur un ensemble de compétences acquises au laboratoire et sur le terrain. L’expérience de la flore locale est indispensable pour apporter une identification de qualité.

Au-delà de donner le nom de l’espèce, le botaniste réunit tous les indices qui lui permettent de rapprocher un individu-arbre rencontré sur le terrain avec une espèce connue et décrite dans la flore de la Nouvelle-Calédonie. Cependant, alors que dans la flore les espèces sont décrites par la totalité de leurs critères morphologiques et anatomiques, le botaniste sur le terrain ne dispose généralement que de quelques informations, qui sont parfois peu accessibles ou uniquement perceptibles aux jumelles. C’est ainsi que si les espèces sont souvent décrites précisément par leurs appareils sexuels (fleurs & fruits), le botaniste de terrain ne peut que rarement compter sur ces critères éphémères pour identifier une espèce; dans une parcelle de forêt moins de 5 % des arbres sont en fleur ou en fruit au moment de leur identification. Le botaniste fait alors appel à des critères qui reposent sur sa propre expertise acquise au fur et à mesure de son observation des plantes à différents âges et dans différents lieux. Cette connaissance lui permet de proposer un nom pour chaque arbre rencontré sur le terrain et s’il est dans l’impossibilité d’atteindre le niveau espèce ou le niveau infra-spécifique, il collecte un rameau de l’arbre qui lui permettra de faire des comparaisons avec une collection d’échantillons archivés dans un herbier. Les botanistes de notre équipe travaillent dans et avec l’herbier de la Nouvelle-Calédonie où ils déposent et identifient les échantillons collectés sur le terrain. Malgré ce travail gigantesque certains arbres ne peuvent pas être identifiés jusqu’au nom de l’espèce, parfois simplement car ils sont trop hauts et inaccessibles sans devoir déployer des moyens démesurés. Ils sont alors exclus des analyses qui portent sur la quantification de la richesse spécifique.
Les inventaires sur le terrain
Les données incluses dans ce site reposent sur la liste des espèces d'arbres de la Nouvelle-Calédonie. Cette liste se base sur les nombreux inventaires de terrain qui s’accumulent dans nos bases de données au fur et à mesure des études que nous réalisons.
Les inventaires recouvrent de nombreuses méthodes mais ils contiennent tous à minima une information relative à la présence d’un arbre dans un lieu donné. Ce couple qui lie l'identification d'une espèce et sa coordonnée géographique constitue une source de donnée fondamentale pour les études écologiques. En effet, la coordonnée permet d'inférer de nombreux paramètres (altitude, milieux, topographie, pluviométrie, substrat, distance aux habitations,...) à l'espèce. Accumulées depuis des années, ces données dessinent progressivement le contour des conditions environnementales propices au développement d’une espèce. Avec ce simple couple de données, nous pouvons dresser les préférences écologiques de chaque taxon en plus de sa distribution dans l’espace géographique.

Les inventaires permettent d’accéder à la notion d’abondance locale en ajoutant le nombre d’individus de chaque espèce comptée sur la superficie effective de l’inventaire. Certaines espèces peuvent alors être rares à l’échelle de la Nouvelle-Calédonie et très abondante localement, comme Acropogon aoupiniensis Morat qui est une espèce micro-endémique qui se rencontre exclusivement aux abords du Mont Aoupinié et qui simultanément est l’espèce la plus abondante de notre parcelle de 1 ha située sur ce massif.
Les inventaires sont également utilisés pour définir les traits fonctionnels d’une espèce, c’est-à-dire ses propriétés intrinsèques qui définissent son comportement. Les traits fonctionnels sont généralement des mesures (diamètre, hauteur, densité de bois, strate, épaisseur et superficie des feuilles,...) qui dressent un profil général pour chaque espèce. Ces profils forment des groupes fonctionnels qui permettent de comparer des peuplements sur la base de leur composition fonctionnelle plutôt que sur leur composition floristique. Par exemple, le diamètre maximal atteint par une espèce dans tous nos inventaires est un trait fonctionnel qui permet de fixer son statut d’arbre puisque seules les espèces qui peuvent atteindre 10 cm de diamètre à 1,3 m du sol (DBH) sont qualifiées d’arbre. Les traits fonctionnels plus complexes à mesurer, tels que les propriétés des feuilles, permettent d’attribuer une résistance de l’espèce aux conditions de l’environnement, notamment l’aridité. Néanmoins, si ces traits fonctionnels sont précieux pour explorer davantage le comportement des espèces, ils ne représentent que des raccourcis dans des processus beaucoup plus complexes que ces traits qualifiés de ‘soft’ ne peuvent appréhender entièrement.
Les analyses statistiques
Parce que la biologie est un domaine où la variation est une propriété fondamentale, il n’est pas possible d’analyser les données sans évaluer la pertinence des résultats en suivant un protocole statistique rigoureux.

En effet, les évidences qui semblent apparaître en consultant rapidement un jeu de données peuvent provenir de différents biais qui concourent à laisser envisager un lien de cause à effet (corrélation), là où seul le hasard est intervenu. Les statistiques sont utilisées pour évaluer la probabilité qu’une hypothèse soit liée au hasard en tenant compte de la taille et de la distribution de l’échantillon. En général ces statistiques sont associées à une hypothèse nulle qui représente une relation liée au hasard. Par exemple, une telle hypothèse pourrait tester si la distribution d’une plante est liée au hasard ou au contraire à l’une ou l’autre des conditions environnementales (substrat, topographie, altitude, milieu,...).
Les statistiques en écologie reposent sur des méthodes très complexes dont la perspicacité pour répondre à la question posée doit toujours être critiquée. Certaines données auto-corrélées, telles que l’altitude et la température qui évoluent ensemble, doivent être traitées pour ne pas sur-évaluer le poids du paramètre seul. Les données utilisées dans ce travail sont traitées en utilisant des méthodes statistiques publiées et validées par un comité de lecture externe. La liste des publications associées à ce travail dévoilent la profondeur des études statistiques et mathématiques qui sont utilisées pour analyser des paramètres qui peuvent paraître simples, comme la distribution de la richesse selon l’altitude (Ibanez et al., 2014; Birnbaum et al., 2015).
Les analyses géographiques

La forêt est une entité qui ne se mesure pas exclusivement par le nombre et la nature de ses espèces. La distribution géographique de cet écosystème permet d’aborder des questions sur son organisation dans le paysage de la Nouvelle-Calédonie. Où se trouve la forêt ? Quelles surfaces occupe-t-elle ? Quel est l’état de sa fragmentation ? sont autant de questions qui relèvent de la géographie. L’étendue de la forêt calédonienne est étudiée au travers des outils les plus modernes, tels que les imageries satellitaires ou aéroportées qui fournissent des données brutes qu’il convient de confronter aux réalités de terrain. La cartographie de la forêt s’appuie sur ces données pour délimiter précisément les ensembles de forêt au travers d’une validation reposant sur une expertise en photo-interprétation associée à un corpus de données. Ces analyses géographiques sont ainsi confrontées aux inventaires de terrain pour paramétrer des modèles qui permettent d’extrapoler une mesure sur toute l’étendue géographique qu’il s’agisse par exemple de la mesure de la biomasse via l’étude de la structure de la canopée sur des images Pléiades (Blanchard et al., 2015, 2016) ou encore de la distribution de la richesse en arbre en utilisant une série d’image Landsat (Pouteau et al., 2018). Bien entendu les analyses statistiques sont utilisées également ici pour définir le domaine de validité de ces modèles.
L’informatique
Toutes ces données ne pourraient être utilisées convenablement sans un stockage rigoureux sous la forme de jeux de données numériques et leur exploitation par des langages informatiques appropriés. Chaque inventaire, propriété fonctionnelle ou taxonomique mentionné dans ce travail est stocké dans une base de données relationnelle, qu’il s’agisse des données textuelles (chiffre, texte) ou géographiques (couche vectorielle). Le stockage, l’analyse et l’exploitation des données sont exclusivement réalisés avec des logiciels dits «open-source» depuis le système d’exploitation (linux) jusqu’aux outils de bases de données (PostgreSQL, PostGIS, Gdal), de cartographie (Qgis), de statistique (R), de développement informatique (Python, Django), de gestion d’images (Gimp, Dartable) ou encore de bureautique (WPS). Le développement de ce site est intégralement conçu et réalisé par notre équipe. Le code est disponible en libre téléchargement sur la plateforme GitHub sous licence GPL-3.0.
Le contenu du site Niamoto reflète l’état de nos bases de données avec une place très réduite pour l'interprétation. C’est ainsi que vous constaterez nos lacunes sur des pans entiers de l’écologie forestière, sur des taxons tellement rares que nous ne les avons jamais rencontrés dans nos inventaires, sur des périodes de phénologie incomplète, sur des contenus déséquilibrés. Cette discipline avance en fonction de ces données, elle progresse continuellement si bien que le contenu de ce site évolue périodiquement pour refléter l'état de nos bases de données. Vous trouverez en bas de ce site, la date de publication associée à la citation du site de Niamoto
Les données
Les données capitalisées dans le projet Niamoto reposent sur de nombreuses publications et collectes acquises dans différents lieux de la Nouvelle-Calédonie. Toutes les données relatives aux arbres et à la forêt proviennent des travaux menés par l’UMR Amap. Néanmoins, nos analyses s’appuient également sur des données externes qui sont utilisées en complément des données biologiques notamment pour étudier la distribution spatiale et environnementale des plantes.
- Le support géographique de la Nouvelle-Calédonie et le catalogue des données téléchargées provient du portail de l'information géographique de la Nouvelle-Calédonie édité par le service de la Géomatique de la Direction des Technologies et des Services de l'Information (georep.nc).
- Les limites administratives (limites provinciales, communales, réserves) sont produites par le gouvernement de la Nouvelle-Calédonie (BD-Topo, DTSI, 2020). Les données sont utilisées conformément à l'Arrêté N°2017-1505/GNC du 04 juillet 2017 et mises à disposition par la province Nord selon les accords de partenariats 12C144 et 16C180 signés respectivement le 27 mars 2013 et le 30 janvier 2018. La commune de Poya, nommée artificiellement Poya Nord, a été divisée selon la limite administrative de la province Nord.
- Les données de pluviométrie sont issues du modèle Aurelhy (Analyse Utilisant le RELief pour les besoins de l'HYdro-météorologie, 1991-2000) produit par Météo-France (Météo-France, 2011).
- Les limites du substrat péridotite (échelle 1:1 000 000), ou substrat ultramafique, sont définies par la DIMENC/SGNC et le BRGM (2010) d’après la carte géologique de Nouvelle-Calédonie (Paris, 1981).
- Le relief est basé sur le modèle SRTM-90 (« Shuttle Radar Topography Mission ») à 90 m de résolution produit par la NASA-NGA (2000) et le Modèle Numérique de Terrain (MNT) à 10 m de résolution crée par la DTSI (2006, révisé en 2012).
- Les limites des concessions minières (échelle 1:10 000) sont issues du Cadastre minier de Nouvelle-Calédonie - SMC/DIMENC - Mise à jour le 08 mars 2013
- Les limites des périmètres de protection des eaux sont issues des travaux des services DAVAR / SDE / MERE (Mesures et Etudes de la Ressource en Eau) révisés le 25 mai 2019. Seuls les Périmètres de Protection Éloignés (PPE) sont considérés dans ce site.